Behavioral Science Essays
The
Attentional Blink
The
Glutamate Theory of Schizophrenia
Non-intelligent
Perception
Anxiety
and the GABA-A receptor subunit
On
The Theory-Ladenness of Observation
ECT in Depressed
Older Adults
Dennett Indented?
The Attentional Blink
Sten M. Andersen
Vision is arguably the most important sense for most human beings; and, as could be expected, several parts of the brain are involved in processing visual information (Bear, Connors, & Paradiso, 2001). As vision is the main provider of information about threats and opportunities in the world, it to a large extent drives behaviour. People are not passive onlookers to life, though, but have expectancies derived from earlier experiences; people have goals, and hence, intentions. These goals and intentions set up a framework for what will be necessary to attend to, and "each of us literally chooses, by his ways of attending to things, what sort of universe he shall appear himself to inhabit" (James, 1890, p. 424). Hence, there is a reciprocal relationship between the available information and what is attended to. It seems like this reciprocal relationship can also be found at a neuronal level. Only about 15% of the input to the lateral geniculate nucleus (LGN) comes from the retina; 80% of the excitatory synapses in the LGN come from the primary visual cortex (Bear, Connors, & Paradiso, 2001). It has been argued that top-down expectations selectively amplify features that are attended to, while at the same time dampening others (e.g., Grossberg, 1998).
Attention seems to be intimately linked to consciousness. We cannot attend to something (be it a physical object or a thought) without at the same time being conscious about the object of attention. Thus, it seems reasonable that the neuronal substrates of attention correlate with those of consciousness. Given that all the information available from the retinas do not enter consciousness, and that that being attended to usually does, it seems like the study of visual attention may give a glimpse into the brain mechanisms of consciousness.
During saccadic eye movement, visual processing is suppressed (Volkman, Riggs, & Moore, 1980). Thus, visual information comes into the perceptual processing system in discrete chunks. To investigate the brain's limitations in processing this information, Raymond, Shapiro, and Arnell (1992) used a task in which letters where presented rapidly to participants, and where participants were asked to identify a target letter and then to indicate whether a probe letter was present or absent in the stream of letters after the target. Raymond, Shapiro, and Arnell found that there was a 270 ms interval starting 180 ms after the target, in which probes were poorly detected. This deficit was not found when a brief blank interval followed the target, or when participants were not asked to identify the target. It was therefore suggested that interference at the time when the letter was being recognised made the system realise there was potential for confusion and uncertainty, and that this led to suppression of visual attention mechanisms; the suppression was dubbed an attentional blink (AB).
A modern, full-blown theory of consciousness needs to be grounded on a solid base of empirical data. The aim of the present study was to investigate the notion of an AB, in an attempt to solidify a small part of the basis for a beginning theory of consciousness. It was hypothesised that there would be a deficit in probe detection until after approximately 450 ms, as this was the extension of the AB as found by Raymond, Shapiro, and Arnell (1992).
Method
Participants
There were 25 participants, all second year psychology students at La Trobe University. All participants were naïve as to the aims and the hypothesis of the experiment.
Materials
A Macintosh computer was used to present the visual stimuli.
Procedure
Participants (P) were seated in front of the computers; all stimuli consisted of letters flashed at the same location of the screen. Distracter letters were black, target letters were red, and the probe was a black 'x'. A stream of letters appeared on the screen, Ps were asked to identify the target letter, and to indicate whether the probe had appeared after the target. The number of distracters varied between trials. An adaptive threshold method was used, such that the time of when Ps could detect the probe 79% of the time could be found.
Results
The data were collated; using an adaptive threshold method, the time of when Ps could detect the probe 79% of the time was found. Mean needed time in milliseconds and standard deviation were calculated. The results are presented in Figure 1.
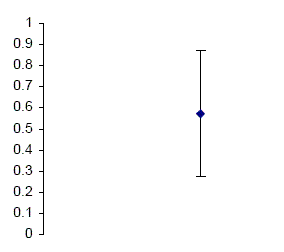
Figure 1. Mean time (ms) needed to perceive probe 79% of the time, and SD.
As illustrated in Figure 1, Ps displayed a refractory period of slightly more
than half a second (M = 571 ms, SD = .298).
Discussion
Consistent with earlier research, the hypothesis that there would be a deficit in probe detection until after approximately 450 ms was supported by the present study.
If the attentional blink is a real mechanism in the human brain, its raison d'être might be speculated upon. As more complex organisms fought for limited resources in a battle for survival, the exact extraction of information from the environment must have become more important. A highly developed perceptual system could thus evolve that would try to minimise uncertainty, as the reduction of uncertainty among a number of alternatives is what constitutes information (Shannon & Weaver, 1964). This could possibly explain a mechanism like the attentional blink (AB); it could be argued that the AB is a way of ensuring that the maximum amount of information is gathered using the limited capacity of the brain.
Using classical information theory in the biological sciences has been fraught with problems. Usually, data has been seen as meaningless until it is interpreted by some understanding being; only then does it become information. But a brain needs to contain information within itself, without there being an external interpreter, or an internal "understander" , a homunculus. This problem seems to have been solved by Edelman and Tononi (2000), who devised a method of measuring information within a network. Interestingly, they showed that for a network to contain information in any real sense, reentrant pathways were needed, just as those found between the lateral geniculate nucleus and the visual cortex.
Even though attention and consciousness seems to be closely related, they should not be equated. For example, even though saccadic eye movements cause discrete chunks of information to be presented to the brain, the images that are perceived appear to be continuous and unbroken. Bregman's (1990) auditory continuity illusion is another striking example. If a steady tone shuts off just as a broadband noise turns on, and then the tone starts again immediately after the noise shuts off, the tone seems to continue through the noise. If the noise is not followed by the tone, however, the tone is not heard through the noise. It is as if the conscious experience of hearing the tone or not hearing the tone is projected backwards in time. Under the condition where the tone reappears after the noise, the perceived tone during noise can be explained in terms of the framework of attention that has been set up in the present paper. The signals in the noise that are similar to the tone are selected by attentional mechanisms, and the other signals are somewhat dampened. Under this condition, attention and consciuousness seems to correlate. When the tone does not reappear after the noise, however, consciuosness of the tone during noise somehow seems to be lost (for further exploration of this theme and a possible explanation, see Grossberg, 1998). Again, this should warn us not to equate attention with consciuosness.
In the present paper, some aspects of the visual system and the mechanisms of attention have been discussed. It has been argued that studying the mechanisms of visual attention might give insights into the neuronal substrates of consciousness. In particular, the phenomenon of an attentional blink (AB) has been looked upon. A reason has been given for why the AB might have evolved, given in terms of maximising information. The concept of information in a brain has been touched upon. Even though attention and consciousness seem to correlate, a warning has been issued not to equate attention with consciousness.
References
| Bear, M. F., Connors, B. W., & Paradiso, M. A. (2001). Neuroscience: Exploring the brain (2nd ed.). US: Lippincott Williams & Wilkins. |
| Bregman, A. S. (1990). Auditory scene analysis: The perceptual organization of sound. Cambridge, MA: MIT Press. |
| Edelman, G. M., & Tononi, G. (2000). Consciousness. How matter becomes imagination. London: Penguin Books. |
Grossberg, S. (1998). The link between brain, learning, attention, and consciousness. Technical Report CAS/CNS-TR-97-018. Boston, MA: Boston University.
| James, W. (1890). Principles of psychology. Vol. 1. New York: Henry Hold and Company. |
| Shannon, C. E., & Weaver, W. (1964). The mathematical theory of communication. Urbana: University of Illinois Press. |
Raymond, J. E, Shapiro, K. L., & Arnell, K. M. (1992). Temporary suppression of visual processing in an RSVP task: An attentional blink? Journal of Experimental Psychology. Human Perception and Performance, 18 (3), 849-60.
Volkman, F. C., Riggs, L. A., & Moore, R. K. (1980). Eyeblinks and visual suppression. Science, 207, 900-2.